

王 璟1,白献晓1,张家庆1,高彬文1,陈俊峰1,高原2,任巧玲1,马 强1,郭红霞1,梁永红1,邢宝松1
(1. 河南省农业科学院 畜牧兽医研究所,河南 郑州450002;2. 新县畜牧局,河南 信阳465550)
摘要:为了研究PPAR信号通路在猪去势诱导的脂肪沉积中的作用,采集去势和非去势淮南猪(共6头,每组各3头)的皮下脂肪组织,提取总RNA后构建文库,利用高通量测序技术检测PPAR通路相关基因表达量,表达量差异基因的筛选阈值为P<0.05且|log2差异倍数|>1。基于mRNA的差异分析以及KEGG富集分析发现,PPAR信号通路中17个基因表达差异达到显著水平,其中14个基因表达量上调、3个基因表达量下调。脑型脂肪酸结合蛋白基因(FABP7)、甘油水孔蛋白基因(AQP7)、视黄素X受体γ基因(RXRγ)等基因显著上调,而磷酸烯醇丙酮酸羧激酶1基因(PCK1)、过氧化物酶体增殖物激活受体γ辅激活因子1α基因(PPARGC1α)、胆固醇7α-羟化酶1基因(CYP
关键词:淮南猪;差异表达基因;高通量测序;PPAR信号通路;脂肪沉积
中图分类号:S828 Q78
Effect of Castration on PPAR Signal Pathway Expression in Huainan Pig Subcutaneous Fat Tissue
WANG Jing1, BAI Xianxiao1, ZHANG Jiaqing1, GAO Binwen1, CHEN Junfeng1, Gao Yuan2, REN Qiaoling1, MA Qiang1, GUO Hongxia1, LIANG Yonghong1, XING Baosong1,
(1. Institute of Animal Husbandry and Veterinary Science, Henan Academy of Agricultural Sciences, Henan province, Zhengzhou 450002, China; 2. Xinxian Bureau of Animal husbandry, Henan province, Xinxian 465550, China)
Abstract: To detect the role of PPAR signal pathway on the castration-induced fat deposition, RNA was isolated from the subcutaneous fat tissues of three castrated Huainan pigs and three intact Huainan pigs. The high-throughput sequencing approach was used to identify mRNA expression difference in castrated and intact pigs. The screening threshold for these genes was P value<0.05 and |log2FoldChange|>1. Based on the difference of the mRNA and the KEGG enrichment analysis, the expression of PPAR signal pathway was the highest in the castrated pigs, of which fourteen genes were upregulated and three genes were downregulated. Lipogenic genes like fatty acid binding protein 7 gene (FABP7), aquaporin 7 gene (AQP7), retinoid X receptor, gamma gene (RXRγ) were upregulated significantly in castrated pigs, and lipolysis genes were downregulated like phosphoenolpyruvate carboxykinase 1 gene (PCK1), peroxisome proliferator-activated receptor gamma, coactivator 1 alpha gene (PPARGC1α) and cytochrome P450, family 7, subfamily A, polypeptide 1 gene (CYP
Key words: Huainan pig; differential expression gene; high throughput sequencing; PPAR signal pathway; fat deposition
中国拥有80多个地方猪品种,列入中国猪种品种志的有48个,是世界上猪种质资源最丰富的国家之一。国外猪种的肉质存在肌内脂肪含量低、肉色灰白、系水力低等缺点,河南地方猪种如淮南猪、确山黑猪等具有肉色鲜红、系水力强、肌内脂肪含量高、肌纤维细等优点,对高档猪肉生产具有重要意义[1]。淮南猪1986年被列入河南省地方优良畜禽品种志,主要产区集中在淮河上游以南,归属于黄淮海黑猪、华北型。该猪种耐热、耐粗饲、产仔数多,生长发育较慢,仔猪出生质量约为0.8 kg,成年公猪体质量平均为149.7 kg,成年母猪为104.9 kg[2]。淮南猪达90 kg的时间大约需要240 d(显著长于国外引进品种),此阶段淮南猪的背膘厚约为3.35 cm,胴体含脂率约为33.5%,均显著高于国外引进品种[3]。因此,淮南猪可以作为研究中国地方猪肉质性状的代表品种。
研究表明,过氧化物酶体增值物激活受体(PPAR)信号通路参与调控脂质代谢、能量代谢、炎症反应、胚胎着床等。PPARs属于核激素受体超家族,核激素受体是一类转录因子,能够结合在DNA并以配体依赖方式调控其转录[4]。PPARs通过结合类视黄醇X受体(RXR)形成异二聚体,结合在靶基因的过氧化物酶体增殖反应元件(PPRE反应元件)上,进而调控其转录水平[5]。PPAR基因家族包含PPARα、PPARβ/δ和PPARγ 3个成员,在多种生物过程中起调控作用,例如能够增强机体胰岛素的敏感性、调节体内糖平衡,尤其是通过调控各种参与细胞脂类代谢基因的表达进而影响脂肪细胞的分化[6]。PPARα基因促进甘油三酯分解为游离脂肪酸,同时刺激肌肉和肝脏β氧化进而降低脂肪酸和甘油三酯的合成[7]。PPARβ能够上调前脂肪细胞中多种成脂相关基因的表达,与细胞的基础脂肪代谢相关,加快能量代谢促进脂肪氧化。PPARγ基因是调控前体脂肪细胞分化的关键转录因子,通过转录水平影响脂肪酸及其衍生物的功能进而调节细胞的增殖分化,其在脂类代谢过程中也起到重要作用[8]。PPAR信号通路其他成员在脂质代谢过程中也起到非常重要的作用,例如过氧化物酶体增殖物激活受体γ辅激活因子1α基因(PPARGC1α)、视黄素X受体γ基因(RXRγ)、脂滴包被蛋白(PLIN1)等。
去势能够促进机体脂肪沉积,在动物生产中广泛应用。仔公猪去势后失去雄性激素分泌的基础,机体出现缺乏雄性激素导致的“肥胖”[9]。开展去势公猪的相关研究有助于阐释动物脂肪沉积的分子机制。目前关于去势增加脂肪沉积有一些研究,但其分子调控机制尚不清楚,因此本研究使用高通量测序技术,对淮南猪皮下脂肪进行测序和生物信息学分析,以期找到PPAR信号通路基因在去势猪和非去势猪皮下脂肪中的表达差异,有助于进一步理解去势促进猪脂肪沉积的分子调控机制。
1 材料和方法
1.1供试动物
选取3对淮南猪仔猪,每对仔猪同窝且出生体质量相似。3对仔猪每对中随机选择1头在5周龄时进行手术阉割,另1头在腹部划相同大小切口作为伪处理,从而产生相同的应激反应。所有猪按照商品猪的饲养流程进行饲养。
1.2 组织样品
于210日龄时对6头试验猪进行屠宰,屠宰后30 min内采集皮下脂肪样品,样品剪成黄豆大小,放在冻存管中立即投入液氮中,运输至实验室后保存于-80 ℃超低温冰箱。
1.3 总RNA提取及质量检测
使用柱式动物组织总RNA抽提纯化试剂盒[生工生物工程(上海)股份有限公司]提取6头猪皮下脂肪的总RNA。通过琼脂糖凝胶电泳检测所提RNA的质量,使用紫外分光光度计(IMPLEN,NanoPhotometer)分析所提RNA的浓度。RNA用干冰包装后,送往北京诺禾致源生物信息科技有限公司测序。
1.4 上机文库制备及质检
检测合格的总RNA构建文库:使用Epicentre Ribo-ZeroTM试剂盒去除核糖体RNA;加入Fragmentation Buffer把mRNA片段化,形成150~200 bp的短片段;以短片段为模板,使用六碱基的随机引物进行第一链cDNA合成,然后加入缓冲液、dNTPs和DNA聚合酶I进行第二链cDNA的合成;利用AMPure XP beads纯化后的双链cDNA进行末端修复、
文库构建完成后,先使用Qubit2.0进行初步定量,稀释文库至1 ng/μL,随后使用Agilent 2100检测文库的插入片段大小(insert size),符合预期后使用qPCR方法对文库的有效浓度进行准确定量(文库有效浓度>2 nmol/L),以保证文库质量。
1.5 高通量测序和生物信息学分析
检测合格的文库按照有效浓度及目标下机数据量的需求进行HiSeq测序。所得原始测序序列,将带接头的、低质量的序列进行过滤,从而得到clean reads。采用Tophat2对过滤后的序列与猪的参考基因组(Scrofa10.2)进行比对分析,得到Mapped Data。基因表达水平用FPKM值表示,是每百万片段中来自某一个基因每千碱基长度的片段数目。通过Mapped Data分析基因的表达差异,筛选阈值为P值<0.05且|log2差异倍数|>1。PPAR信号通路主要成员信息见表1。
表1 PPAR信号通路部分成员介绍
Tab.1 The introduction of partial factors in PPAR signal pathway
|
英文名 English name |
中文名 Chinese name |
功能 Function |
相关疾病 Disorders |
参考文献 Reference |
|
PPARGC1α |
过氧化物酶体增殖物激活受体γ辅激活因子1α |
线粒体增殖和呼吸;脂肪形成和分化;肝糖异生;促进棕色脂肪组织形成;肌纤维转化 |
胰岛素抵抗;胰岛素敏感性;代谢紊乱;II型糖尿病;肥胖 |
[10] |
|
PCK1 |
磷酸烯醇丙酮酸羧激酶1 |
葡糖异生关键酶;甘油异生 |
胰岛素抵抗;II型糖尿病;脂肪肝 |
[11] |
|
CYP |
胆固醇7α-羟化酶1 |
胆汁酸合成限速酶;脂肪沉积;糖代谢;能量动态平衡;炎症反应 |
胆汁淤积;高胆固醇血症;吸收障碍;动脉粥样硬化 |
[12] |
|
FABP7 |
脑型脂肪酸结合蛋白7 |
脂肪酸摄取和胞内运输;细胞增殖;T细胞自我平衡 |
恶性胶质瘤;肾癌;乳腺癌 |
[13] |
|
AQP7 |
水通道蛋白7 |
甘油分解及转运;精子分化成熟;机体能量平衡和葡萄糖稳态 |
胰岛素抵抗;高血糖症;肥胖 |
[14] |
|
RXRγ |
视黄素X受体γ |
脂肪细胞分化;脂质代谢;胆固醇代谢;甾醇和脂肪酸代谢平衡 |
高脂血症;肿瘤 |
[15] |
|
PPARδ |
过氧化物酶体增殖物激活受体δ |
细胞增殖和分化;脂肪酸氧化;自适应产热 |
血脂异常;胰岛素敏感性;胰岛素抵抗;代谢紊乱;肥胖 |
[16] |
|
ACSL1 |
长链脂酰辅酶A合成酶1 |
长链脂酰CoA合成酶;能量代谢;脂肪沉积 |
胰岛素抵抗;肥胖 |
[17] |
|
OLR1 |
氧化型低密度脂蛋白受体1 |
甘油三酯储存;脂质代谢;肾损伤;脂肪细胞分化 |
高脂血症;动脉粥状硬化;粥样硬化斑块;心血管疾病 |
[18] |
|
ACOX2 |
三羟基粪甾烷酰CoA氧化酶2 |
参与长支链脂肪酸和胆汁酸的降解;重度精神发育迟滞 |
zellweger综合征 |
[19] |
|
SORBS1 |
Sorbin和SH3结构域蛋白1 |
刺激胰岛素发挥功能 |
胰岛素抵抗;肥胖 |
[20] |
|
PPARα |
过氧化物酶体增殖物激活受体α |
脂质代谢;脂肪酸合成和氧化;糖异生;生酮作用 |
酒精性心肌病;血脂异常;胰岛素抵抗;胰岛素敏感性;脂肪肝;动脉粥样硬化 |
[21] |
|
PLIN2 |
脂滴包被蛋白 |
脂肪分解调控的“分子开关” |
粥样硬化斑块;胰岛素敏感性;胰岛素抵抗;肥胖 |
[22] |
|
PPARγ |
过氧化物酶体增殖物激活受体γ |
脂肪细胞分化;葡萄糖稳态 |
动脉粥样硬化;胰岛素抵抗;胰岛素敏感性;II型糖尿病;肥胖 |
[23] |
|
UBC |
泛素 |
蛋白质降解;DNA修复;细胞周期调控;激酶修饰 |
乳腺肿瘤 |
[24] |
|
PPARGC1β |
过氧化物酶体增殖物激活受体γ辅激活因子1β |
脂肪氧化;葡萄糖代谢 |
肥胖 |
[25] |
2 结果与分析
2.1差异表达基因分析
图1所示为去势与非去势猪的皮下脂肪组织差异表达mRNA KEGG富集表达聚类,其中代谢通路相关基因表达量较高用深颜色表示,表达量较低用浅颜色表示。所列出的20个通路中,有与机体糖脂代谢调控相关的如II型糖尿病通路(14个差异基因)、胰岛素信号通路(28个差异基因)、AMPK信号通路(29个差异基因)、甘油酯代谢通路(16个差异基因)等。已知PPAR信号通路相关基因调控机体能量代谢,尤其与胰岛素抵抗、II型糖尿病、肥胖等糖脂代谢紊乱疾病相关[6]。本研究中PPAR信号通路在去势组颜色较深而非去势组颜色较浅,说明其表达量在去势组表达量较高而非去势组较低。与其他通路相关基因表达量相比,去势组PPAR信号通路相关基因表达量非常高,提示该通路在去势猪脂肪沉积过程中起重要作用,值得进一步深入研究。
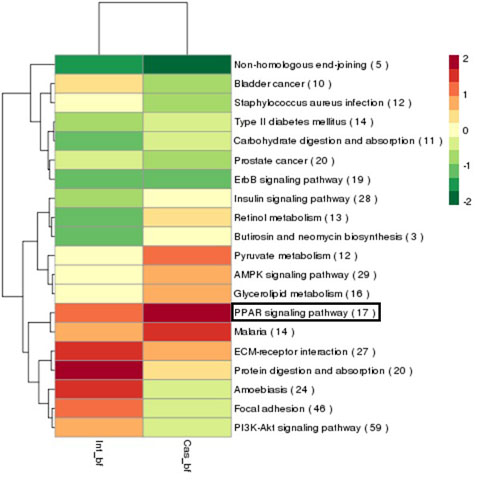
图1. 差异表达mRNA KEGG富集表达聚类
Fig. 1 The KEGG of differently expressed mRNA
2.2 PPAR通路相关基因差异表达情况
由表2可知,与非去势组相比,去势组PPAR信号通路共有17个基因表达量变化达到筛选阈值。其中上调基因14个,下调基因3个。去势组FPKM值变化范围为0.069~547.697,而非去势组为0.292~152.724。差异表达基因|log2差异倍数|的范围从-4.568到4.123,平均值为2.012。
表2 去势组与非去势组PPAR通路相关基因表达量
Tab. 2 The expression of PPAR signal pathway genes in castrated and intact pigs
|
名称 |
去势猪 FPKM |
非去势猪 FPKM |
|log2差异倍数| |
P值 |
上调/下调 | |||
|
Name |
Castrated pig FPKM |
Intact pig FPKM |
|log2Fold Change| |
P value |
Up/Down | |||
|
FABP7 |
5.095 |
0.292 |
4.123 |
0.000 |
上调 |
| ||
|
AQP7 |
77.414 |
16.020 |
2.273 |
0.000 |
上调 |
| ||
|
RXRγ |
14.970 |
3.146 |
2.251 |
0.000 |
上调 |
| ||
|
PPARδ |
24.621 |
5.303 |
2.215 |
0.000 |
上调 |
| ||
|
ACSL1 |
547.697 |
128.246 |
2.094 |
0.001 |
上调 |
| ||
|
PLIN2 |
10.524 |
2.822 |
1.899 |
0.000 |
上调 |
| ||
|
OLR1 |
3.614 |
0.996 |
1.859 |
0.002 |
上调 |
| ||
|
ACOX2 |
7.855 |
2.495 |
1.655 |
0.001 |
上调 |
| ||
|
SORBS1 |
83.219 |
30.609 |
1.443 |
0.002 |
上调 |
| ||
|
PPARα |
22.404 |
8.541 |
1.391 |
0.003 |
上调 |
| ||
|
PLIN1 |
391.687 |
152.724 |
1.359 |
0.005 |
上调 |
| ||
|
PPARγ |
22.472 |
9.485 |
1.244 |
0.008 |
上调 |
| ||
|
UBC |
1.558 |
0.682 |
1.192 |
0.132 |
上调 |
| ||
|
PPARGC1β |
1.662 |
0.754 |
1.140 |
0.020 |
上调 |
| ||
|
PCK1 |
17.437 |
39.269 |
-1.171 |
0.005 |
下调 |
| ||
|
PPARGC1α |
0.069 |
0.344 |
-2.319 |
0.019 |
下调 |
| ||
|
CYP |
0.082 |
1.934 |
-4.568 |
1.000 |
下调 |
| ||
3 讨论与结论
FABP7基因上调倍数最高,其|log2差异倍数|为4.123,该基因最早是从脑组织中分离出来的,在肝脏中表达量也较高,其主要作用是调节脂肪酸的摄取和细胞内运输。吴丹[26]通过全基因组关联分析发现FABP7基因与北京油鸡体质量及屠体性状相关。方心灵等[27]研究发现,FABP7基因在岭南黄羽肉公鸡禁食48h显著下调。何侃等[28]研究发现,FABP7基因与SREBP
AQP7基因属于甘油水孔蛋白亚类,位于脂肪细胞细胞膜上,当机体需要能量的时候,其主要将甘油从脂肪细胞内转运至血液,其在脂肪代谢中具有关键的媒介作用[29]。本研究中,AQP7基因在两个组差异倍数仅次于FABP7,提示猪去势后其雄性激素分泌减少能够调控AQP7基因的表达水平。但雄性激素是直接调控FABP7还是通过转化为雌激素间接调控?其具体调控机制还需进一步开展研究证明。
CYP
PPARGC1α是PPARγ的核转录辅激活因子,其主要在线粒体含量丰富的组织中表达,例如棕色脂肪组织和肌肉组织,而在白色脂肪组织中表达量降低[34],所以本研究结果中PPARGC1α的FPKM值较低,尤其在去势组其FPKM值仅为0.069,为PPAR信号通路中最低值。PPARGC1α通过与不同转录因子结合参与适应性产热、线粒体的生物合成、肌纤维转换、脂肪酸氧化等消耗能量的生物过程,本研究中与非去势猪相比,去势猪脂肪组织的分解代谢降低而合成代谢增高,所以该基因在非去势组表达量更高。而PCK1(磷酸烯醇式丙酮酸羧激酶)基因,其编码的PEPCK酶不仅是糖异生过程的限速酶,还与甘油异生密切相关[35]。有研究发现,PCK1基因表达量增加导致肝脏葡萄糖合成增加,出现胰岛素抵抗等II型糖尿病症状,说明该基因在葡萄糖代谢稳态具有重要作用[36] 。小鼠的肝脏特异性敲除PCK1基因后出现脂肪肝[37],说明该基因与脂肪沉积呈负相关,这与本研究结果一致,脂肪沉积增多的去势猪PCK1基因表达量显著低于非去势组。
综上所述,猪去势后出现雄性激素缺乏导致的脂肪沉积增多,即脂肪的合成代谢大于分解代谢。在这个过程中,PPAR信号通路共有17个基因表达量发生显著变化,其中13个上调,3个下调,差异基因|log2差异倍数|最大值为4.568,平均值为2.012,提示该信号通路在猪去势肥育过程中起到关键作用。下一步研究需要明确雄性激素如何影响这些基因的表达。
参考文献:
[1]卜鸿静,岳磊,韵晓冬,等.引进猪种与我国地方猪种生产效率及肉质特性比较[J].山西农业科学,2014,42(1):74-77.
[2]豆成林.淮南猪肉品质特性研究[D].杨凌:西北农林科技大学,2008.
[3]豆成林,周枫.淮南猪、豫南黑猪和长白猪氨基酸组成及含量的分析与比较[J].食品工业,2013,34(9):200-202.
[4]吕妍,徐金娥.PPAR与糖尿病脂代谢的相关性[J].齐鲁医学杂志,2013,28(3):277-279.
[5]熊敏.PPARs调控家禽脂肪代谢的基因网络研究[D].武汉:华中农业大学,2011.
[6]柳晓峰,李辉.PPAR基因与脂肪代谢调控[J].遗传,2006,28(2):243-248.
[7]陈雪香,刘合焜.PPARα基因多态性与2型糖尿病关系研究[J].医学综述,2008,14(6):897-899.
[8]蒋金航,马云,王新庄.PPARγ基因调控脂肪细胞分化的研究进展[J].中国畜牧杂志,2014,50(9):91-95.
[9]蔡兆伟,潘永明,陈亮,等.睾酮缺乏对高脂饮食小型猪血脂和肝内脂质沉积的影响[J].中国比较医学杂志,2015,25(1):40-44, 45.
[10] 李密杰,张春梅,蓝贤勇,等.共激活因子PGC-1α的研究进展[J].中国牛业科学,2012,38(4):35-38.
[11] Saigi-Morgui N, Vandenberghe F, Delacretaz A,et al. Association of PCK1 with body mass index and other metabolic features in patients with psychotropic treatments[J]. Journal of Clinical Psychopharmacology, 2015, 35(5): 544-552.
[12] Bao L D, Li C Q, Peng R,et al. Correlation between the decrease of cholesterol efflux from macrophages in patients with type II diabetes mellitus and down-regulated CYP7A1 expression[J]. Genetics and Molecular Research : GMR, 2015, 14(3): 8716-8724.
[13] 熊胜.鹅Lb-FABP基因克隆及填饲对鹅Lb-FABP、Apo-B和Apo-A1基因表达的影响[D].南京:南京农业大学,2013.
[14] 潘虹,杨东英,陈宏.水-甘油通道蛋白7基因(AQP7)的研究进展[J].中国牛业科学,2012,38(5):53-57.
[15] 黄萌,许尚忠,昝林森,等.牛RXRG基因cDNA的克隆及生物信息学分析[J].西北农林科技大学学报(自然科学版),2008,36(11):1-5, 10.
[16] 梁晶,王红卫,程利安,等.北京黑猪PPARD基因G32E多态位点与脂肪沉积相关性状的关联分析[J].中国畜牧兽医,2015,42(7):1793-1799.
[17] Joseph R, Poschmann J, Sukarieh R,et al. ACSL1 is associated with fetal programming of insulin sensitivity and cellular lipid content[J].Molecular Endocrinology, 2015, 29(6): 909-920.
[18] Khaidakov M, Mitra S, Kang B Y,et al. Oxidized LDL receptor 1 (OLR1) as a possible Link between obesity, dyslipidemia and cancer[J]. PLoS One, 2011, 6(5): e20277, 1-9.
[19] Johansson A, Curran J E, Johnson M P,et al. Identification of ACOX2 as a shared genetic risk factor for preeclampsia and cardiovascular disease[J].European Journal of Human Genetics, 2011, 19(7): 796-800.
[20] Germain M, Pezzolesi M G, Sandholm N A,et al. SORBS1 gene, a new candidate for diabetic nephropathy: Results from a multi-stage genome-wide association study in patients with type 1 diabetes[J]. Diabetologia, 2015, 58(3): 543-548.
[21] Vernia S, Cavanagh-Kyros J, Garcia-Haro L A,et al. The PPAR alpha-FGF21 hormone axis contributes to metabolic regulation by the hepatic JNK signaling pathway[J]. Cell Metabolism, 2014, 20(3): 512-525.
[22] Kozusko K, Tsang V H,Bottomley W,et al. Clinical and molecular characterization of a novel PLIN1 frameshift mutation identified in patients with familial partial lipodystrophy[J]. Diabetes, 2015, 64(1): 299-310.
[23] Wu Y R, Cun Y N, Jing Dong,et al. Polymorphisms in PPARD, PPARG and APM1 associated with four types of Traditional Chinese Medicine constitutions[J].遗传学报, 2010(06): 371-379.
[24] 周鹏,张子平,王艺磊,等.大黄鱼ubc9基因的克隆和组织表达[J].生物技术通报,2009(8):76-82.
[25] 高晓娟,吉红,常志光, 等.PGC-1β和SREBP-1c在猪脂肪细胞分化过程中的相互作用[J]. 中国生物化学与分子生物学报,2011(06):533-539.
[26] 吴丹.北京油鸡体重和屠体性状的全基因组关联研究[D].北京:中国农业科学院,2012.
[27] 方心灵,束刚,王松波,等.禁食及恢复采食状态下鸡能量代谢及下丘脑转录谱的比较[J].畜牧与兽医,2012,44(S1):143.
[28] 何侃,蒋越,王起山,等.基于芯片数据分析关于SREBP-1c调控脂肪代谢过程作用机理的研究[J].黑龙江畜牧兽医,2010,23(13):1-4.
[29] Lebeck J. Metabolic impact of the glycerol channels AQP7 and AQP9 in adipose tissue and liver[J]. Journal of Molecular Endocrinology, 2014, 52(2): R165-R178.
[30] 迟静,翟成凯,郭延波,等.CYP7A1基因多态性对脂代谢异常人群影响[J].中国公共卫生,2013,29(4):491-493.
[31] 李万贵,张依裕,潘兰兵,等.鸭CYP7A1基因编码区C129T突变对肉质和脂肪性状的遗传效应分析[J].广东农业科学,2012,39(24):158-160.
[32] 张依裕,李万贵,王单单.番鸭CYP7A1基因多态性对肉质和血清生化指标的效应研究[J].中国畜牧杂志,2015,51(11):5-9.
[33] 邵丹,王来娣,张蕊,等.鹅肥肝形成相关基因的研究进展[J].中国畜牧兽医,2012,39(9):42-46.
[34] 尚进,靳烨.PPARGC1α基因与肉质品质相关性研究进展[J].肉类研究,2012,26(3):42-44.
[35] 张泽宾.PCK1转录调控机制及功能性分子标记鉴定[D].石河子:石河子大学, 2013.
[36] 李剑虹,杜晓彤,刘火,等: 冷刺激下AA肉鸡PCK1基因的表达量[J]. 贵州农业科学 2014(08):142-146.
[37] 董艳,张宏利,冯绮文,等.人PCK1基因克隆及其表达载体的构建[J]. 上海交通大学学报(医学版) 2006(02):135-137.